Mở đầu
Lúa gạo (Oryza sativa L.) là một trong những loại lương thực chính quan trọng nhất trên thế giới, cũng là loài quan trọng để nghiên cứu di truyền chức năng thực vật [1]. Tuy nhiên, cải thiện năng suất lúa gạo vẫn luôn là vấn đề cấp bách, do sự gia tăng liên tục của dân số thế giới, sự suy thoái của môi trường và giảm diện tích đất canh tác. Năng suất hạt gạo bao gồm 3 yếu tố chính, bao gồm số bông hữu hiệu trên cây, số hạt trên bông và khối lượng hạt. Khối lượng hạt bị ảnh hưởng bởi hình dạng hạt, kích thước hạt (chiều dài, chiều rộng và độ dày), độ đầy của nhân [2, 3]. Kích thước hạt là một đặc điểm nông học quan trọng quyết định tiềm năng năng suất có thể được cải thiện bằng cách sử dụng kỹ thuật di truyền [1, 4]. Trong những năm gần đây, nhiều phương pháp có yếu tố di truyền liên quan đến năng suất của hạt, bao gồm các yếu tố liên quan đến các con đường tín hiệu như protein G, ubiquitin-proteasome, hormone, các yếu tố điều hòa phiên mã... đã được thực hiện [5, 6]. Trong đó, các nghiên cứu gần đây đã khám phá ra một số yếu tố phiên mã hoạt động như những chất điều chỉnh kích thước hạt chính, cho thấy nhiều hứa hẹn trong việc cải thiện và nâng cao năng suất cây lúa [7].
Yếu tố phiên mã
Yếu tố phiên mã là các protein tham gia vào quá trình chuyển đổi phiên mã DNA thành RNA. Các yếu tố phiên mã bao gồm một số lượng lớn các protein (không bao gồm RNA polymerase), bắt đầu và điều chỉnh quá trình phiên mã của các gen [8]. Các yếu tố phiên mã có thể hoạt động đơn lẻ hoặc tương tác với các protein khác trong một phức hợp nhằm kích hoạt hoặc kìm hãm RNA polymerase bám vào vùng khởi động phiên mã để phiên mã thông tin di truyền từ DNA sang RNA của các gen cụ thể [9]. Hoạt động của các yếu tố phiên mã cho phép biểu hiện duy nhất từng gen trong các loại tế bào khác nhau và trong quá trình phát triển.
Các yếu tố phiên mã cải thiện kích thước hạt lúa
Các yếu tố phiên mã điều hòa sự biểu hiện gen có thể ảnh hưởng rộng rãi đến sự phát triển kích thước hạt, do đó quyết định năng suất của lúa [10]. Chẳng hạn, gen GLW7 mã hóa yếu tố phiên mã SPL13, giúp cải thiện kích thước hạt bằng cách tăng sự mở rộng tế bào glume trong vỏ trấu, giúp tăng chiều dài hạt và khối lượng hạt [11]. Gen GW8 mã hóa yếu tố phiên mã SPL16, làm tăng chiều rộng và năng suất của hạt bằng cách thúc đẩy quá trình phân chia tế bào và làm đầy hạt [12]. Tương tự, gen GS2 mã hóa bộ điều hòa phiên mã OsGRF4, điều chỉnh kích thước hạt bằng cách thúc đẩy phân chia tế bào và mở rộng tế bào trong vỏ trấu [13]. Trong khi đó, gen qLGY3 mã hóa yếu tố phiên mã miền MADS1 được liên kết với các hạt dài và mảnh do sự gia tăng phân chia tế bào theo hướng dọc ở lớp biểu bì bên ngoài [14]. Gen GL4 mã hóa cho yếu tố phiên mã MYB và kiểm soát chiều dài hạt bằng cách điều chỉnh sự kéo dài tế bào theo chiều dọc ở lớp men bên ngoài và bên trong ở gạo châu Phi [15]. Sự tăng cường biểu hiện của yếu tố phiên mã HLH107 làm tăng kích thước hạt bằng cách tăng số lượng tế bào theo chiều dọc của vỏ trấu [16]. Nghiên cứu mới đây nhất chỉ ra rằng, sự biểu hiện tăng cường của yếu tố phiên mã DREB1C ở lúa cho năng suất cao hơn 41,3-68,3% so với cây thông thường, do số hạt trên bông và khối lượng hạt tăng lên. Khả năng quang hợp được nâng cao, đồng thời tăng các chất đồng hóa quang hợp dẫn đến sự thúc đẩy tăng trưởng hoạt động vận chuyển và hấp thụ nitơ tăng cao làm tăng năng suất, đặc biệt là trong điều kiện nitơ thấp. Sự biểu hiện tăng cường của yếu tố phiên mã DREB1C còn kích thích cây ra hoa sớm hơn và tích lũy sinh khối cao hơn ở giai đoạn đầu so với cây đối chứng trong điều kiện dài ngày [17].
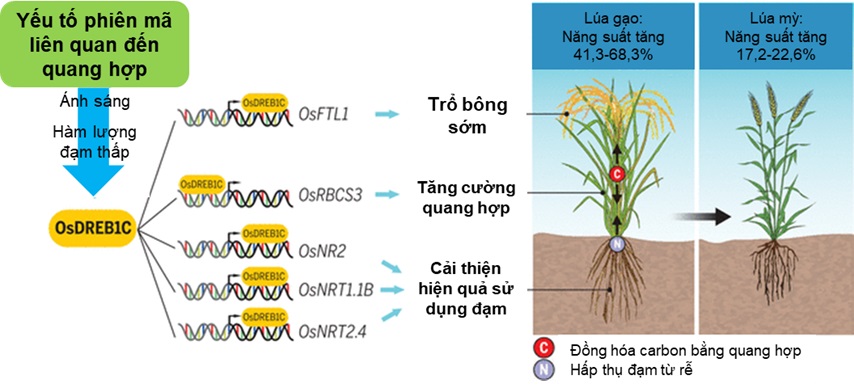
Yếu tố phiên mã giúp tăng năng suất và rút ngắn thời gian sinh trưởng của cây lúa.
OsDREB1C được xác định có khả năng đáp ứng với ánh sáng và lượng nitơ thấp gồm 118 yếu tố phiên mã liên quan đến quá trình quang hợp. OsDREB1C kích hoạt sự phiên mã của nhiều gen mục tiêu, giúp tăng cường quang hợp, cải thiện việc sử dụng nitơ và ra hoa sớm, làm tăng năng suất đáng kể ở lúa và lúa mỳ.
Ngược lại, một số yếu tố phiên mã được phát hiện có vai trò ức chế sự phát triển hạt. Yếu tố phiên mã GBP1 ngăn chặn chiều dài hạt, trái ngược với chức năng tăng cường độ dài hạt của yếu tố phiên mã GBP3 [18]. Bên cạnh sự hạn chế bởi vỏ gai, sự phát triển nội nhũ hoặc sự lấp đầy của hạt cũng rất quan trọng để quyết định kích thước hạt. Yếu tố phiên mã MADS29 điều chỉnh sự suy thoái của các mô mẹ, do đó làm đầy hạt [19]. Sự biểu hiện quá mức hoặc ức chế biểu hiện của yếu tố phiên mã PIL15 trong nội nhũ làm cho kích thước, trọng lượng hạt gạo giảm và tăng tương ứng [1]. Sự biểu hiện của yếu tố phiên mã NF-YB1 làm chậm đáng kể sự phát triển của nội nhũ, dẫn đến hạt nhỏ [20]. Như vậy, các yếu tố phiên mã không chỉ có vai trò thúc đẩy, mà còn kìm hãm sự phát triển của vỏ trấu, sự lấp đầy nội nhũ hạt, gây ảnh hưởng trực tiếp đến kích thước và khối lượng của hạt. Hiểu được các vai trò này, có thể giúp các kỹ thuật di truyền tác động đến sự tăng cường năng suất lúa thông qua cải thiện kích thước hạt gạo.
Yếu tố phiên mã kiểm soát số hạt trên bông lúa (GNP)
Số hạt trên bông là một đặc điểm nông học quan trọng góp phần vào năng suất lúa. Số hạt trên bông chủ yếu được xác định bởi các nhánh bông và sự hình thành cành [21]. Yếu tố phiên mã SPL9 được phát hiện có vai trò quan trọng trong việc hình thành và phát triển các nhánh hoa thứ cấp ở lúa. SPL9 được biểu hiện cao trong các bông non đang phát triển sớm, phù hợp với chức năng điều chỉnh sự phát triển của cấu trúc bông lúa. Đột biến SPL9 làm giảm đáng kể các nhánh thứ cấp và số lượng hạt trên bông [21].
Nghiên cứu mới đây [22] chỉ ra rằng, yếu tố phiên mã RGN1 có ảnh hưởng đến số lượng hạt và cấu trúc hoa lúa. Đột biến gen RGN1 gây ra sự thiếu hụt hạt ở các cành thứ cấp. Trong khi đó, một alen có lợi là RGN1C có nguồn gốc từ lúa hoang dã lại làm cho bông dài hơn. Việc phát hiện và xác định RGN1 cung cấp cơ sở lý thuyết để hiểu cơ chế phân tử của sự hình thành hạt lúa; đồng thời RGN1 sẽ là nguồn gen quan trọng để lai tạo phân tử giúp năng suất cao hơn [22].
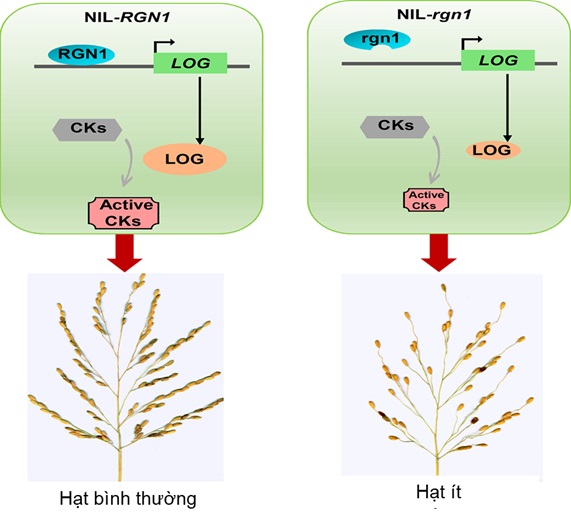
Mô hình minh họa vai trò của yếu tố phiên mã RGN1 đối với cấu trúc bông lúa.
Yếu tố phiên mã RGN1 bám vào vùng khởi động của gen LOG để kích hoạt sự biểu hiện của LOG. LOG biến đổi các cytokinin từ dạng không hoạt động (inactive CKs) thành các dạng hoạt động (active CKs) để khuếch đại đường truyền tín hiệu cytokinin quy định sự phát triển một bông hoa bình thường. Nếu protein RGN1 không liên kết được với vùng khởi động của LOG sẽ ngăn cản sự khuếch đại tín hiệu cytokinin khiến các nhánh hoa phát triển hình chùy lỏng lẻo và thưa hạt.
Một gen điều hòa REM20 (REPRODUCTIVE MERISTEM 20) mã hóa yếu tố phiên mã B3 chưa được công nhận trước đây kiểm soát GNP ở lúa thông qua sự thay đổi các trình tự lặp lại ngược (IR) trong trình tự khởi động của nó, làm thay đổi mức độ biểu hiện của nó và góp phần vào sự biến đổi GNP giữa các giống lúa. Nghiên cứu chỉ ra rằng, các biến thể di truyền mới đóng vai trò quan trọng cho sự đa dạng GNP ở lúa, tiết lộ cơ chế phân tử cơ bản trong việc điều chỉnh biểu hiện gen quan trọng về mặt nông học và cung cấp một chiến lược đầy hứa hẹn để cải thiện sản lượng lúa bằng cách điều khiển trình tự IR chứa yếu tố điều hòa [23].
Năng suất lúa thường bị giới hạn bởi sự cân bằng giữa tổng số hạt trên bông và số nhánh. IPA1 là một gen đa hình điển hình ở cây lúa làm tăng số hạt trên bông nhưng làm giảm đẻ nhánh, do có vùng điều hòa phiên mã là vị trí mục tiêu cho yếu tố phiên mã An-1 để ngăn chặn biểu hiện IPA1 ở bông và rễ. Khi vùng điều hòa phiên mã của IPA1 bị xóa, sẽ giải quyết sự cân bằng giữa số hạt trên bông và số nhánh, dẫn đến năng suất hạt được nâng cao đáng kể ở mỗi cây [24].
*
* *
Yếu tố phiên mã không chỉ tham gia ảnh hưởng đến kích thước và trọng lượng của hạt, mà còn tham gia tích cực vào sự hình thành và phát triển cấu trúc bông lúa, yếu tố quyết định số hạt trên bông. Hầu hết các yếu tố phiên mã có vai trò thúc đẩy sự hình thành và phát triển của hạt cũng như cấu trúc bông lúa, một số khác lại đóng vai trò kìm hãm nhằm duy trì sự cân bằng tự nhiên giữa số lượng hạt và cấu trúc của bông lúa. Cải thiện di truyền cây trồng đòi hỏi phải cân bằng những sự cân bằng phức tạp gây ra bởi sự đa dạng gen và sự liên kết với các yếu tố phiên mã kiểm soát, điều hòa biểu hiện của các gen. Do đó, nhắm mục tiêu đến các vùng điều hòa gen sẽ giúp phân tích các tác động cân bằng và cung cấp nguồn mục tiêu phong phú để lai tạo các tính trạng bổ sung có lợi, nhằm tăng năng suất và chất lượng cho cây lúa.
TÀI LIỆU THAM KHẢO
[1] X. Ji, et al. (2019), "The basic helix-loop-helix transcription factor, OsPIL15, regulates grain size via directly targeting a purine permease gene OsPUP7 in rice", Plant Biotechnology Journal, 17(8), pp.1527-1537.
[2] Y. Xing, Q. Zhang (2010), "Genetic and molecular bases of rice yield", Annual Review of Plant Biology, 61(1), pp.421-442.
[3] D. Zeng, et al. (2017), "Rational design of high-yield and superior-quality rice", Nature Plants, 3(4), p.17031.
[4] K. Century, et al. (2008), "Regulating the regulators: The future prospects for transcription-factor-based agricultural biotechnology products", Plant Physiology, 147(1), pp.20-29.
[5] J. Zuo, J. Li (2014), "Molecular genetic dissection of quantitative trait loci regulating rice grain size", Annual Review of Genetics, 48(1), pp.99-118.
[6] N. Li, R. Xu, Y. Li (2019), "Molecular networks of seed size control in plants", Annual Review of Plant Biology, 70(1), pp.435-463.
[7] A. Pereira, et al. (2014), "Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress"", Nature Communications, 5(1), p.5302.
[8] D.S. Latchman (1997), "Transcription factors: An overview", The International Journal of Biochemistry & Cell Biology, 29(12), pp.1305-1312.
[9] E. Eliopoulos, et al. (2020), "Transcription factors and evolution: An integral part of gene expression (Review)", World Academy of Sciences Journal, 2(1), pp.3-8, DOI: 10.3892/wasj.2020.32.
[10] V.F. Irish, et al. (2008), "An Arabidopsis F-box protein acts as a transcriptional co-factor to regulate floral development", Development, 135(7), pp.1235-1245.
[11] B. Han, et al. (2016), "OsSPL13 controls grain size in cultivated rice", Nature Genetics, 48(4), pp.447-456.
[12] X. Fu, et al. (2015), "The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality", Nature Genetics, 47(8), pp.949-954.
[13] Q. Qian, et al. (2015), "A rare allele of GS2 enhances grain size and grain yield in rice", Molecular Plant, 8(10), pp.1455-1465.
[14] X. Fu, et al. (2018), "G-protein βγ subunits determine grain size through interaction with MADS-domain transcription factors in rice", Nature Communications, 9(1), p.852.
[15] Z. Zhu, et al. (2017), "A single-nucleotide polymorphism causes smaller grain size and loss of seed shattering during African rice domestication", Nature Plants, 3(6), p.17064.
[16] J. Wan, et al. (2018), "Overexpression of OsbHLH107, a member of the basic helix-loop-helix transcription factor family, enhances grain size in rice (Oryza sativa L.)", Rice, 11(41), DOI:10.1186/s12284-018-0237-y.
[17] W. Zhou, et al. (2022), "A transcriptional regulator that boosts grain yields and shortens the growth duration of rice", Science, 377(6604), DOI:10.1126/science.abi8455.
[18] S. Yu, et al. (2018), "Divergent functions of the GAGA-binding transcription factor family in rice", The Plant Journal, 94(1), pp.32-47.
[19] L.L. Yin, H.W. Xue (2012), "The MADS29 transcription factor regulates the degradation of the nucellus and the nucellar projection during rice seed development", The Plant cell, 24(3), pp.1049-1065.
[20] J.J. Xu, X.F. Zhang, H.W. Xue (2016), "Rice aleurone layer specific OsNF-YB1 regulates grain filling and endosperm development by interacting with an ERF transcription factor", Journal of Experimental Botany, 67(22), pp.6399-6411.
[21] B. Tu (2021), "OsSPL9 Regulates grain number and grain yield in rice", Front Plant Sci., 12, p.682018.
[22] Z. Li (2022), "RGN1 controls grain number and shapes panicle architecture in rice", Plant Biotechnology Journal, 20(1), pp.158-167.
[23] Y. Wang (2021), "Enhancing rice grain production by manipulating the naturally evolved cis-regulatory element-containing inverted repeat sequence of OsREM20", Molecular Plant, 14(6), pp.997-1011.
[24] https://www.nature.com/articles/s41587-022-01281-7.